
1 Introduction
Wastewater salinity is a noteworthy problem in biological treatment processes all over the world. Very large amounts of high saline wastewater (mainly NaCl) are generated in tanning, petroleum, chemical production, food processing as well as textile industry (Lefebvre and Moletta, 2006; Kishor et al., 2021). Also, in many coastal areas, seawater is used to flush toilets to alleviate the shortage of freshwater, which could lead to increased salinity in wastewater treatment plants (WWTPs) (Liu et al., 2019; Li et al., 2022).
Biological treatment processes, especially activated sludge systems, are widely used in the removal of pollutants on account of their low cost and high efficiency. But high salinity could cause poor treatment performance of biological WWTPs, since high osmotic pressure is lethal to microbes and can inhibit the enzyme activity (Duan et al., 2015; Maqbool et al., 2018; Han et al., 2021; Luo et al., 2021). Previous researches showed that salinity reduced the diversity of bacterial community, altered the compositions of nitrifiers and denitrifiers, and further inhibited the activity of nitrifying bacteria (Luo et al., 2015; Zhao et al., 2016; Wang et al., 2017a; García-Ruiz et al., 2018; Wang et al., 2020a). To date, the activated sludge microbial community has been investigated extensively, but seldom focuses on the microbial interactions among microbial taxa during system operation.
Activated sludge system is a complex micro-ecosystem, in which various bacterial taxa interact with each other through energy transfer and exchange of matter and information to form a large, complex ecological network (Yuan et al., 2021). Through such network connections, the biological treatment process can achieve desirable performance in removal of organic matters and nutrients (Wang et al., 2019; Ya et al., 2021). For example, there is a symbiotic relationship between ammonia oxidizing bacteria (AOB) and nitrite oxidizing bacteria (NOB), as AOB provides nitrite for NOB as the metabolic substrate, while NOB removes nitrite to reduce its inhibitory effect on AOB. For another example, heterotrophic Saprospiraceae can hydrolyze protein to small molecular amino acids, which can be utilized by other heterotrophic bacteria as metabolic substrates (Xia et al., 2008). Other typical microbial cooperative interactions in activated sludge systems include: 1) AOB and NOB can provide substrates such as nitrite and nitrate for denitrifying bacteria (Teske et al., 1994; Baumann et al., 1997; Mota et al., 2005); 2) CO2 released by heterotrophic bacteria can be assimilated by autotrophic AOB and NOB (Peng et al., 2015); and 3) soluble microbial products released after cell death and the lysis of AOB and NOB can be used as organic carbon source for denitrifying bacteria (Miao et al., 2016). It is expected that some physicochemical changes (e.g. influent salinity) might disturb the microbial interactions among various functional populations (e.g., heterotrophic bacteria, nitrifiers, denitrifiers, polyphosphate accumulating organisms (PAO), etc.), and accordingly impact the performance efficiency in activated sludge systems (Wang et al., 2017b; Wang et al., 2019). Hence, understanding the response of microbial interactions to salinity perturbation is significant to develop operational strategies to resist unfavorable disturbance and further improve the nitrification-denitrification processes of activated sludge systems. However, information regarding microbial interactions and how they respond to elevated salinity has been rarely reported. Recently, by performing a novel RMT-based network analysis, the ecological interactions among members of the community can be assessed and keystone populations of the complex community can be identified, providing a novel insight into the dynamic changes of microbial interaction under physicochemical changes (Sun et al., 2020; Yuan et al., 2021).
In this study, the potential relationship between system performance and microbial interaction dynamics was evaluated and the keystone species were identified by using network analysis in activated sludge microbial communities during a salinity gradient of 0%–3%. A dynamic variation of molecular ecological networks (MENs) was detected to explore the following questions: 1) How does the overall network structure respond to elevated salinity? 2) How does the subnetwork structure of different phylogenetic taxa respond to elevated salinity? 3) How do the functional bacteria and keystone species respond to elevated salinity?
2 Materials and methods
2.1 Reactor construction and operational strategy
A sequencing biological reactor (SBR) (Fig. S1) with 7.7 L effective volume was operated at 25 ± 1 °C for 80 d. Seed sludge was activated sludge that had been domesticated by a low C/N ratio environment. The synthetic wastewater contained 400 mg/L COD, 80 mg/L NH4+-N, 4 mg/L TP, 36 mg/L KCl, 14 mg/L CaCl2·2H2O, 90 mg/L MgSO4·7H2O and 0.3 mL/L trace elements solution. The composition of trace elements solution was according to Smolders et al. (1994). The system was continuously exposed to 0%, 1%, 2%, and 3% salinities (using NaCl), named phases A, B, C and D, respectively, and each phase was conducted for 20 d. The above-mentioned reagents were analytical reagents and purchased from Tianjin Guangfu Technology Development Co., Ltd., China.
The reactor ran for 6 h in one cycle, pumping wastewater for 10 min, stirring for 1.5 h, aeration for 1.5 h, stirring for 2 h, settling for 0.5 h, drainage for 10 min, and idle stage for 10 min. The rotation speed in the stirring stage was 200 (± 20) r/min, and the aeration volume in the aeration stage was 1.6 (± 0.3) L/min. And the wastewater intake and drainage were 4 L per cycle. MLSS was maintained at 3000 (± 500) mg/L.
2.2 Analytical methods
COD, NH4+-N, NO2−-N, and NO3−-N levels were measured every three days. NH4+-N from aerobic-stage start, NO2−-N and NO3−-N from aerobic-stage end were measured every three days. NH4+-N, NO2−-N, and NO3−-N in a cycle at the end of each phase were measured every 30 min. Total nitrogen (TN) was the sum of the concentrations of NH4+-N, NO2−-N, and NO3−-N. pH and dissolved oxygen (DO) were measured every day. MLSS and MLVSS were measured every 10 d. COD, NH4+-N, NO2−-N, and NO3−-N were detected in accordance with Standard Method (Wang et al., 2017b).
2.3 Microbial DNA extraction and high throughput sequencing
A total of 32 activated sludge samples were generated from four phases during the final eight consecutive days of each phase. DNA extraction was performed using the FastDNA SPIN Kit for Soil (MP Biotechnology, France) following the manufacturer’s recommendations (Hu et al., 2021). 16S rRNA gene amplicon sequencing was performed on Illumina MiSeq PE300 platform in Majorbio Co., Ltd. (Shanghai, China) by using the primers 338F and 806R for bacteria in the V3–V4 hypervariable region. Then raw data were quality-filtered according to our previous literature (Wang et al., 2017b). Operational taxonomic units (OTUs) were obtained by using UPARSE (v11) with 97% sequence similarity. And then the OTUs were taxonomically annotated using RDP Classifier (v2.13) against the 16S rRNA database (Silva v138) with a confidence threshold of 0.7. Raw sequences generated in this paper have been deposited in NCBI SRA under the accession number PRJNA732228.
2.4 Data analysis and network construction
Sobs, Shannon, Shannoneven indices represented the diversity of the bacterial community. The statistical analysis was proposed in R (v.3.5.1). In addition, four MENs (named 0%, 1%, 2%, and 3% networks) were constructed to reveal the possible co-occurrence patterns among species by Molecular Ecological Network Analysis Pipeline. The detailed methods of the construction of experimental and random networks were shown in Text S1, and iDIRECT (Inference of Direct and Indirect Relationships with Effective Copula-based Transitivity) was applied to disentangle direct from indirect relationships in the constructed networks according to Xiao et al. (2022) in Text S2. Gephi 0.9.2 and Cytoscape 3.6.1 were used for visualization of the networks.
3 Results and discussion
3.1 System performance
As shown in Fig.1(a), the salinity gradient (1%–3%) had no obvious impact on organic matter removal, as the COD removal efficiency was always higher than 94% in all phases. In comparison, although the effluent TN was less than 0.93 mg/L at lower salinity (1% and 2%), the TN concentration suddenly increased to 14.93 mg/L under 3% salinity. Fig.1(b) showed the concentrations of NH4+-N, NO2−-N, and NO3−-N in a cycle at 0%–3% salinities. NH4+-N remained at a relatively high level (14.64 mg/L) after O-stage when exposed to 3% salinity, while the concentrations of NO2−-N and NO3−-N were very low and even couldn’t be detected at the end of the anoxic stage, which revealed that nitrification was more severely inhibited than denitrification under high salinity. Fig.1(c) further illustrated that NO3−-N was the main nitrification product after O-stage under 0% salinity, and the NAR (the accumulation rate of nitrous nitrogen: NO2−-N/(NO2−-N + NO3−-N)) was less than 0.23%. While, the NAR increased to 99.72% along with the salinity from 1% to 3%, and NO3−-N was undetectable after O-stage, suggesting NO2−-N was the main product of nitrification. These results demonstrated shortcut nitrification-denitrification was achieved under salt conditions, which was consistent with previous research (Ye et al., 2009). In addition, the production of NO2−-N after O-stage declined in the presence of 3% salinity, indicating higher salinity also inhibited the shortcut nitrification.

Fig.1 Dynamic changes of nitrogenous substances in SBR along a salinity gradient. Removal efficiencies of COD and TN (a); the concentrations of NH4+-N, NO2−-N, and NO3−-N in one cycle (b); dynamic changes of nitrogenous substances before and after O-stage (c); under different salinity. A-stage, O-stage and A-stage in a cycle represented anaerobic stage, aerobic stage and anoxic stage, respectively. |
3.2 Community structure
All samples were rarefied to 33264 sequences for subsequent comparative analyses. A total of 891 bacterial OTUs were determined based on 97% sequence similarity level. The rarefaction curves were exhibited in Fig. S2(a). Sobs and Shannon indices were both significantly negatively correlated (P < 0.01) with elevated salinity (Fig. S3), implying that salinity could reduce the abundance and diversity of bacterial communities. Besides, high salinity (3%) greatly altered the bacterial community compositions (Fig. S2). For example, α-Proteobacteria (59.49%) was the dominant population under 0%–2% salinity, while β-Proteobacteria (70.17%) became the most abundant class in the present of 3% salinity. The above results showed that salinity had a significant influence on microbial diversity and composition.
3.3 Characteristics of association networks
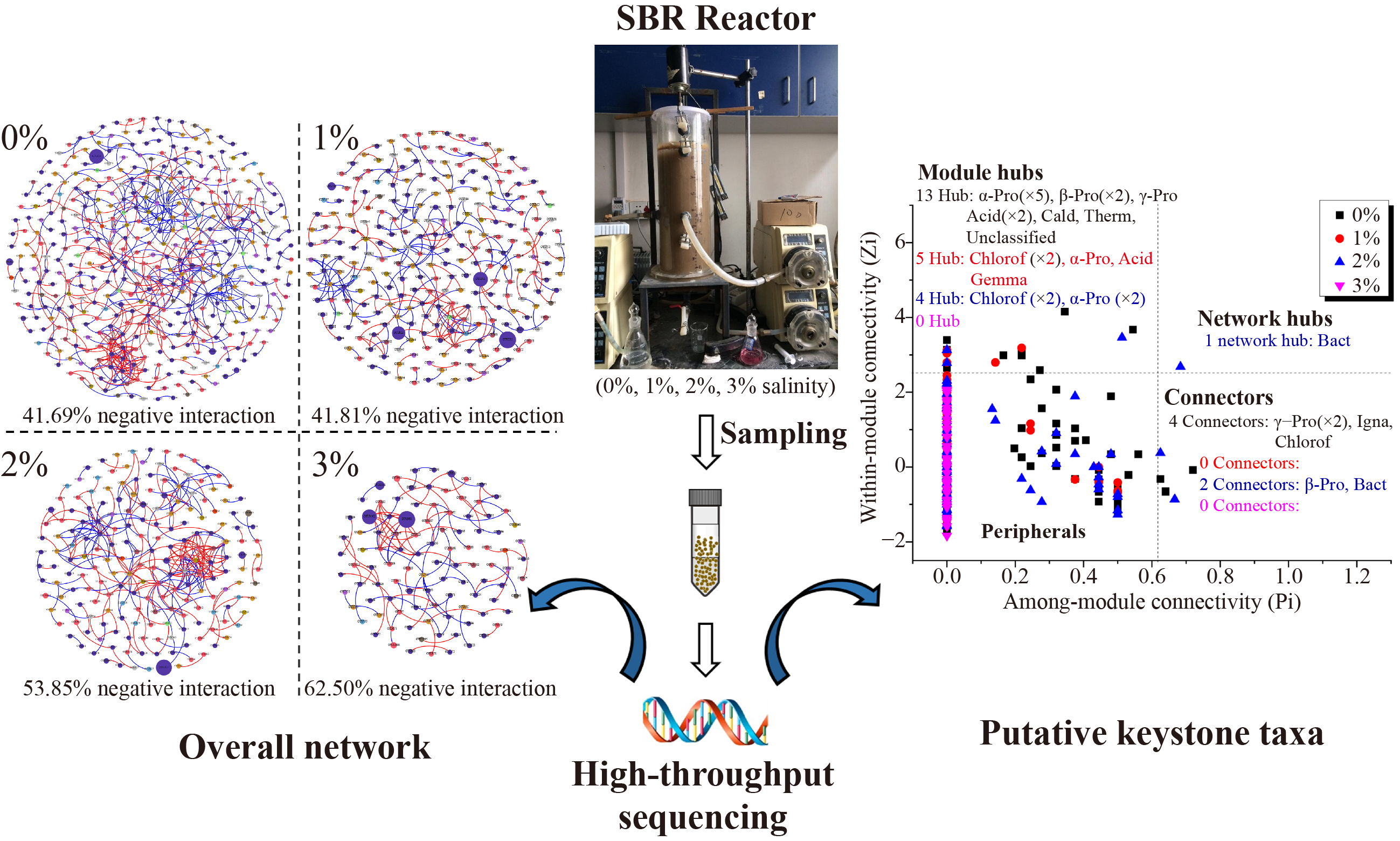
To explore the effect of elevated salinity on microbe-microbe interactions in an activated sludge system, we constructed four bacterial networks for each salinity and removed spurious/indirect links by using iDIRECT method (Fig.2). The network topological features of original and iDIRECT-processed networks were exhibited in Table S1. A considerable portion of the links (23.18%, 22.64%, 30.95%, and 38.80%, corresponding to 0%–3% salinity) was removed in elevated salinity networks. As a consequence, the average clustering coefficient (avgCC) significantly decreased under 0%–3% salinity compared to the original networks, while the modularity of the iDIRECT-derived networks increased significantly compared to the original networks. Subsequent analyses were based on the networks generated by iDIRECT. Besides, the iDIRECT-derived bacterial networks significantly (P < 0.05) differed from corresponding randomized networks, suggesting the empirical network structures were nonrandom. The topology of all networks fit the power-law distribution very well (R2 from 0.802 to 0.961), and the topological features were higher than those in corresponding random networks. These metrics indicated that these four networks clearly exhibited small world, scale-free, and modularity properties (Girvan and Newman, 2002; Blonder et al., 2012; Deng et al., 2012).

3.4 Effects of elevated salinity on overall networks
Although the identical threshold (St) 0.92 was adopted to construct networks, these four microbial networks differed profoundly (Fig.2). Higher salinity formed smaller networks with fewer nodes, as there were 407, 260, 196, and 102 nodes with 590, 287, 299, and 112 links across four salinity elevated networks. The average degree (avgK) and avgCC decreased at 1% salinity, and then significantly increased under higher salinities, while the modularity presented an opposite trend (Fig.3 and Table S1). Among these indexes, avgK represents the microbial interaction complexity, and avgCC indicates the connectivity between a node and its neighbors. The average path distance (GD) not only describes the shortest path between two nodes, but also represents the exchange speed of substances and information between two nodes (Luo et al., 2006). The variations of network topological indexes indicated 1% salinity weakened the connection between nodes. However, when exposed to higher salinity (2% or 3% salinity), the increase in avgK and avgCC, as well as the decrease in GD led to more complex and tighter network connection, suggesting the intense communications among microorganisms under higher salinities.

In microbial networks, organic matter and nutrient removal depend on various types of interactions, which generally fall into two categories, positive connection and negative connection. Positive relationships mainly include mutualism, while negative relationships are generally due to competition, as predation and amensalism are unlikely to emerge in activated sludge systems (Deng et al., 2016; Wu et al., 2016; Ya et al., 2021). In this study, the proportion of negative links increased from 41.69% to 41.81%, 53.85%, and 62.50% when exposed to elevated salinity from 0% to 3% (Fig.3). The highest proportion of competitive interactions under 3% salinity might lead to the deterioration of TN removal. In addition, the higher salinities (2% and 3%) also reduced the modularity, as the module numbers of four networks (0%, 1%, 2%, and 3% networks) were 55, 47, 33, and 22 with modularity values of 0.837, 0.894, 0.782 and 0.786. Modularity, describing how well a community could be divided into modules, is regarded as a topological indicator and is crucial for system stability and resilience (Olesen et al., 2007; Wu et al., 2016). The module could be considered as a functional unit since the OTUs within one module might tend to own similar ecological niches (Luo et al., 2006; Eiler et al., 2012). It appeared that 2%–3% salinity decreased the modules (i.e., functional units) and this could be another potential reason causing the poor performance of nitrogen removal.
3.5 Effects of elevated salinity on subnetworks
The subnetworks of major phyla containing the top seven nodes with the highest connectivity were generated in Fig.4. Proteobacteria was the dominant phylum in the SBR system, and in the subnetworks of Proteobacteria, the number of nodes declined from 62 to 46, 45, and 18, respectively, and the proportions of negative relationship were 70.71%, 38.78%, 45.31%, and 78.79%, respectively, under elevated salinity process. It appeared that higher salinity formed a smaller size network and the most intensive competition was found in 3% salinity. Proteobacteria mainly contributed to the removal of organic matter and nutrients in the activated sludge system, therefore, the minimum network size and the most intensive competition probably led to the deterioration performance at 3% salinity condition. Within Bacteroidetes, the subnetwork size became bigger at 2% salinity and then turned smaller at 3% salinity, as there were 35, 33, 41, and 21 nodes according to 0%–3% salinities. For Chloroflexi, 3% salinity also dramatically reduced the size and interactions, while promoted the competitive relationships in the corresponding subnetworks. The smallest network size and complexity, as well as the most intensive competition among these major phyla under 3% salinity probably led to the deterioration of system performance.

Although higher salinity levels reduced the community diversity and simplified the network connection, functional organisms could still form interactive networks in high salinity environments. Therefore, we further discussed the potential effects of the salinity gradient on some functional bacteria subnetworks. For example, Nitrosomonas and Nitrosomonadaceae were two typical AOB, while their response to elevated salinity was different. The relative abundance of Nitrosomonas was 0.02%, 0.03%, 0.05% and 1.55%, while Nitrosomonadaceae was 1.29%, 0.47%, 0.10% and undetected with salinity increasing from 0% to 3%. Based on the changes in abundance, Nitrosomonadaceae was gradually replaced by Nitrosomonas, which might make a greater contribution to nitrification under higher salinity (especially 3%) in the activated sludge system. Consistently, 1, 0, 2, and 4 OTUs belonged to Nitrosomonas, and 7, 3, 1, and 0 OTUs belonged to Nitrosomonadaceae were detected in the salinity elevated networks. These OTUs and their first neighbors were used to construct the subnetworks of Nitrosomonas and Nitrosomonadaceae (Fig.5). At the salinity gradient of 0%–2%, the percentage of negative links decreased from 80.00% to 66.67% in the Nitrosomonas subnetworks and also reduced from 40.00% to 0.00% in Nitrosomonadaceae subnetworks. When salinity increased to 3%, the negative link proportions rapidly rose to 93.73% in Nitrosomonas subnetwork, and no nodes related to Nitrosomonadaceae were detected. It suggested that higher salinity completely inhibited the relationship between Nitrosomonadaceae and other populations, and it also started to suppress Nitrosomonas simultaneously, leading to the decline of NO2−-N production after O-stage under 3% salinity shown in Fig.1(b). In addition, Nitrospira, as the representative NOB, was only found in 0% network, which manifested that salinity seriously inhibited the nitrification of NO2−-N resulting in the elevation of NAR over 99.72% under 1%–3% salinity in activated sludge system.

Some denitrifiers, such as Acinetobacter, Thauera, Azoarcus, Paracoccus, Denitromonas, and Defluviicoccus, were also used to construct subnetworks (Fig.5). The size and complexity of subnetworks of Acinetobacter and Thauera were both declined by elevating salinity. For example, the four Acinetobacter subnetworks contained 17, 14, 12, and 8 nodes with 14, 11, 9, and 5 links, respectively, along with a salinity gradient. Besides, the elevated salinity stress decreased the abundance of Acinetobacter (from 2.10% to 0.06%–0.61%), whereas enhanced Thauera abundance (from 1.70% to 2.66%–11.18%). Although a similar trend of these two subnetworks was observed, the abundance trends of Acinetobacter and Thauera were different. This inconsistency highlighted the necessity of studying the bacterial interaction, as it provided a pivotal dimension to understand the microbial assembly process rather than the simple community richness and diversity. In addition, the most complex subnetwork of Azoarcus appeared at 2% salinity, and the most complex subnetwork of Paracoccus appeared at 3% salinity, while the most complex subnetwork of Denitromonas appeared at 0% salinity. Defluviicoccus, an endogenous denitrifier, was found dominant at 0%–2% salinity (relative abundance: 37.15%, 52.07%, 35.01%, and 8.80% according to 0%–3% salinity) under this intermittent aeration sewage treatment system (Wang et al., 2020b). However, it appeared that 1%–3% salinity seriously reduced the microbial interactions in Defluviicoccus subnetworks, as the subnetworks contained 74, 32, 44, and 14 nodes with 63, 25, 41, and 10 links, respectively (Fig. S4). It is worth noting that the percentage of negative relationships in subnetworks of Acinetobacter, Thauera, Azoarcus, Paracoccus, and Denitromonas reduced from 92.86%, 88.24%, 75.00%, 100%, and 87.50% to 40%, 50%, 71.43%, 33.33%, and 84.62%, respectively, according to 0% and 3% salinities, indicating denitrifiers established more proportion of cooperative relationships with other bacteria to resist 3% salinity stress. The above results indicated that elevated salinity could have different influences on the microbial interactions of various bacteria with the same function to ensure the stability of the activated sludge system, even at 3% salinity.
3.6 Putative keystone taxa of association networks
In addition to the above-mentioned functional microorganisms, topological keystone species also play irreplaceable roles in ecological networks. The topological role of these species could be defined by Zi-Pi parameter (Fig.6(a)). In this study, over 97.12% of the nodes were classified as peripherals, while only 22 module hubs and six connectors as well as only one network hub were identified in the elevated salinity networks (Tables S2). Among these 22 module hubs, 11 derived from Proteobacteria, 6 belonged to Chloroflexi, 3 related to Actinobacteria, 1 related to Gemmatimonadetes, and 1 belonged to unclassified bacteria, all of which merely amounted to very low abundance ranging from 0.01% to 1.79% (Fig.6(b)). Six connectors from four networks were derived from four phyla, 50.00% of connectors were Proteobacteria, and the others were derived from Bacteroidetes, Chloroflexi, and Ignavibacteriae. Similarly, all connectors also presented relatively low abundance (ranging from 0.01% to 0.37%) (Fig.6(c)). Only one OTU (OTU 959) was identified as the network hub at 2% salinity in this study, and this is the only few times that network hubs were identified by network analysis (Wu et al., 2016; Ya et al., 2021). Previous studies have suggested that connectors and hubs could be regarded as key species, as they play over-proportional ecological role in maintaining community structure and function stability than some abundant species (Olesen et al., 2007; Shi et al., 2016). Among these key species, eight module hubs (OTUs 28, 1075, 599, 198, 581, 663, 750, and 771) as well as one connector (OTU 1005) were closely related to Defluviicoccus, Thauera, norank_f__Blastocatellaceae_Subgroup_4, norank_o__JG30-KF-CM45, norank_f__Gemmatimonadaceae, Gemmobacter, and Azoarcus, all of which were denitrifiers (Kämpfer et al., 2015; Wang et al., 2020b; Wang et al., 2020c; Huang et al., 2022). Hub OTU 955 as well as connector OTU 635, derived from the Candidatus Competibacter, were considered as denitrifying glycogen accumulating organisms (DGAO) (Xu et al., 2020). Besides, two module hubs (OTUs 252 and 736) and one connector (OTU 704) attributed to the family Caldilineaceae, as well as one connector OTU 898 attributed to Pseudoxanthomonas were capable of degrading lignocellulose (Zhang et al., 2019; Tang et al., 2020). Two hubs (OTUs 793 and 744) belonged to the family Anaerolineaceae. Members of Anaerolineaceae were found abundant in anaerobic system and were responsible for the removal of carbohydrates and cell tissues (Miao et al., 2016). Connector of OTU 555 was related to order Ignavibacteriales, and they were key players in the observed shift from denitrification to dissimilatory reduction of nitrate to ammonium (Grießmeier et al., 2017). Furthermore, two module hubs (OTUs 117 and 259) attributed to Acidobacteria and one connector OTU 380 attributed to Bacteroidetes were found to degrade some complicated organic substances. OTU 629 was related to order SC-I-84 and OTUs 601, 43 and 577 belonged to unclassified bacteria, and their specific functions remain unknown.

Interestingly, three nodes (OTUs 28, 771, and 736) served as module hubs in different networks. OTU 28 functioned in 0% and 2% networks, and OTU 771 functioned in 1% and 2% networks, and OTU 736 functioned in 0% and 1% networks. The other 89.66% of hubs and connectors were unique. No putative keystone species were found in 3% network. Notably, OTU 28 was derived from Defluviicoccus and considered as an endogenous denitrifier as well as a glycogen accumulating organism (GAO), and OTU 771 was also responsible for denitrification. OTUs 28 and 771 and their first neighbors were presented in Fig. S5. There were 21, 18, 14, and 3 nodes with 20, 20, 24, and 2 links corresponding to 100% positive correlations with salinity increasing, which suggested 1%–2% salinity reduced the subnetwork size but promoted the complexity and collaboration between populations to resist the high salinity, however, OTUs no longer served as key taxa and more species could not be detected under 3% salinity. Although 0%–2% salinities greatly altered the mostly putative key microbial populations, various species could successionally perform the same function under elevated salinity. For example, OTUs 1075, 599, 955,198, 581, 663, 635 from 0% network, OTUs 750, 771 from 1% network and OTUs 28, 1005 from 2% network all functioned as denitrifiers. Besides, OTUs 117, 736, 704, 898 from 0% network, OTUs 252, 259 from 1% network, and OTUs 793, 794, 380 from 2% network were all responsible for degrading organic substances. Hence, functional redundancy might be a potential factor for the keystone taxa dynamics with the salinity increasing. Specially, the relatively low abundances of these putative keystone species in this study indicated that some low-abundant microorganisms could play a disproportionately more significant role than abundant species in maintaining microbial community stability in activated sludge systems.
The above network analysis could identify that the putative keystone species were critical in maintaining the community structure and functional stability of the activated sludge system in response to a salinity gradient, although they were dynamic and less abundant. Therefore, network analysis has much potential for exploring these keystone populations interactions to gain a deeper understanding of the critical roles of these species.
4 Conclusions
The mechanism underlying the succession of ecological network is often overlooked in microbial ecology. Here, we evaluated the effects of elevated salinity on ecological networks of activated sludge system by performing a novel RMT-based network analysis. Our results found 3% salinity inhibited TN removal and reduced the diversity of microbial communities. Network analysis revealed that overall networks under higher salinity conditions (2% and 3%) exhibited more complex, tighter networks, and more competitive bacterial interactions. Subnetworks of bacteria with same function (such as AOB, NOB, and denitrifiers) differed substantially when exposed to elevated salinity. The connection between Nitrospira (NOB) and other species was seriously inhibited under 1%–3% salinity resulting in the elevation of NAR over 99.72%. In addition, keystone species (hubs and connectors) were dynamics when exposed to different salinity and played crucial roles in maintaining system stability, despite the low abundances in microbial community. Collectively, this work improved our understanding of the relationship between system performance and microbial interaction dynamics of activated sludge microbial communities in response to elevated salinity. Further studies are necessary to verify the salinity effects in more parallel reactors with control.